13.5.2. Продолговатый мозг и варолиев мост
Продолговатый мозг и варолиев мост вместе составляют задний мозг. Анатомически и функционально задний мозг связан со спинным мозгом, мозжечком и большими полушариями головного мозга. В нем замыкаются дуги многих сложнокоординирован-ных двигательных процессов.
Сегментарное строение, четко выраженное в спинном мозге, стерто в продолговатом. Сюда вступают афферентные волокна от слуховых рецепторов, рецепторов полости рта, кожи лицевой части головы, сердца, крупных сосудов и легких, слизистых оболочек глаз, ряда рецепторов желудка, печени, поджелудочной железы, тонких кишок. Но в органах грудной и брюшной полости имеются также рецепторы, от которых афферентные волокна идут не в продолговатый, а в спинной мозг. Эфферентные волокна нейронов продолговатого мозга иннервируют все мышцы лица, сердце, бронхи, гортань, пищевод, желудок, поджелудочную железу, печень, кишечник. Иннервация внутренних органов осуществляется парасимпатическими волокнами совместно с симпатическими волокнами, лежащими в спинном мозге.
Из области продолговатого мозга и варолиева моста выходят следующие черепно-мозговые нервы: тройничный, отводящий, лицевой, слуховой, языкоглоточный, блуждающий, добавочный, подъязычный.
Продолговатый мозг играет большую роль в регуляции мышечного тонуса. Скелетные мышцы находятся в постоянном тонусе, за счет которого преодолевается действие силы тяжести и тело сохраняет равновесие. Тонус мышц имеет рефлекторную природу. Начало дуги этого рефлекса лежит в мышцах и сухожилиях.
Если головной мозг вместе со средним мозгом и его красными ядрами удален, а продолговатый мозг сохранен и не отделен от спинного, то у млекопитающих развивается сильное, часами и сутками удерживаемое напряжение разгибательных мышц (рис. 13.13).
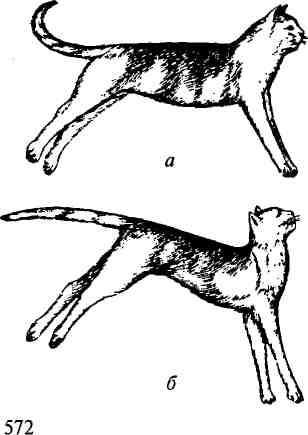
Конечности такого животного вытянуты, и согнуть их можно лишь с трудом; шейные мышцы, разгибающие голову, напряжены; позвоночник несколько выгнут дугой кверху.
Таким образом, все мышцы, напряжение которых противостоит действию силы тяжести, оказываются после отделения продолговатого мозга от среднего в состоянии длительного тонического напряжения. Это
Рис. 13.13. Децеребрационная ригидность:
а — кошка с выключенными большими полушариями (путем перевязки сосудов); б— то же при одновременном выключении мозжечка
резкое усиление тонуса разгибательных мышц носит название децеребрационной ригидности (децеребрация — удаление мозга выше продолговатого). Тонус разгибателей при децеребрационной ригидности превышает тонус сгибателей, но длительное напряжение сгибателей способствует тому, что конечности становятся как бы твердыми опорами и могут удерживать тяжесть тела.
Децеребрационная ригидность не наступает, если средний мозг животного связан с продолговатым мозгом, а также если продолговатый отделен от спинного мозга. Отсюда следует, что в продолговатом мозге лежат нервные клетки, которые могут приводить в возбуждение спинальные нейроны, иннервирующие разгибатели. Если продолговатый мозг связан со средним мозгом, ригидность также не наступает, т. е. от среднего мозга исходят влияния, угнетающие стимуляцию разгибательных рефлексов со стороны продолговатого мозга.
Заключительный этап в возникновении непомерного тонуса связан с рефлекторной деятельностью ядра вестибулярного нерва. Волокна от рецепторов вестибулярного аппарата в продолговатом мозгу контактируют с массой клеток, образующих так называемое вестибулярное ядро. Оперативное удаление вестибулярного ядра и особенно его части, известной как ядро Дейтерса, резко ослабляет децеребрационную ригидность. Разрушение вместе с ядром Дейтерса ретикулярной формации продолговатого мозга совершенно ее прекращает.
Для развития децеребрационной ригидности необходимо поступление в ЦНС импульсов от рецепторов разгибательных мышц. Инъекция в эти мышцы раствора новокаина или перерезка соответствующих задних корешков снимает тонус мышц-разгибателей. Следовательно, децеребрационную ригидность надо рассматривать как результат усиления рефлексов, осуществляемых совместно спинным и продолговатым мозгом в ответ на импульсы, поступающие как от рецепторов мышц-разгибателей, растягиваемых под действием силы тяжести, так и от рецепторов лабиринта. Эти рефлексы в норме всегда уравновешиваются воздействиями, рефлекторно возникающими в среднем мозге, мозжечке, а у высших млекопитающих главным образом в коре больших полушарий.
Роль продолговатого мозга в осуществлении децеребрационной ригидности обусловлена постоянным поступлением импульсов от важнейших рецепторов дыхательной и кровеносной систем, а также с лабиринтов, вследствие этого он постоянно воздействует на рефлекторные центры спинного мозга, повышая их возбудимость. При отделении спинного мозга от продолговатого возбудимость нервного центра никогда не достигает той степени, при которой тонус мышц-разгибателей в ответ на импульсы от их рецепторов рефлекторно усиливался бы до степени, характеризующей децеребрационную ригидность.
Децеребрационная ригидность — это состояние, конечно, совершенно нефизиологичное, но по ней можно судить о взаимоотношениях между разными образованиями ЦНС. Надо заметить, что механизм этого процесса еще не совсем ясен. Это состояние не всегда наступает после децеребрации, иногда оно выражено очень резко, а иногда почти не проявляется.
Продолговатый мозг участвует в регуляции проводниковых рефлексов и в осуществлении таких актов, как ходьба и стояние, для которых необходима интегративная деятельность ЦНС. Буль-барное животное, у которого сохранены лишь спинной и продолговатый мозг, способно к более сложным рефлекторным процессам, чем спинальное, так как все основные функции в более полной мере объединены и скоординированы.
Варолиев мост является центральной частью заднего мозга. Он имеет вид толстого белого вала. Серое вещество образуют ядра черепно-мозговых нервов (V—VIII пары), ядра ретикулярной формации и собственные ядра моста.
Белое вещество состоит из продольных и поперечных пучков нервных волокон. Продольные волокна идут по нижней поверхности моста в продолговатый мозг, образуя в нем пирамиды. Сверху они составляют восходящие и нисходящие пути двусторонних связей моста. Поперечные волокна направляются в ножки мозжечка.
Чувствительные волокна черепно-мозговых нервов проводят сигналы от вкусовых рецепторов языка, рецепторов кожи и мышц головы, глаз и зубов. Эфферентные волокна обеспечивают мимику. Таким образом, рефлекторная деятельность варолиева моста дополняет защитные и пищевые рефлексы, осуществляемые продолговатым мозгом.
Источник
Продолговатый мозг
Непосредственным продолжением спинного мозга у всех позвоночных животных является продолговатый мозг, формирующийся в процессе онтогенеза из заднего мозгового пузыря головного конца мозговой трубки. Продолговатый мозг сохраняет в чертах своего строения отдельные признаки сегментарных примитивных отношений, типичных для спинного мозга. Однако правильность в распределении серого и белого вещества здесь значительно нарушена. В результате структурных и функциональных перестроек, сопровождающих процесс филогенеза, скопления клеточных тел привели к образованию т.н. ядер продолговатого мозга, являющихся центрами более или менее сложных рефлекторных функций. Часть этих ядер входит в состав ретикулярной формации продолговатого мозга. Из 12 пар черепно-мозговых нервов восемь (с 5 по 12) берут свое начало от продолговатого мозга. Многочисленные волокна этих нервов (соматические и вегетативные) образуют связи между продолговатым мозгом и самыми разнообразными органами.
В продолговатом мозге находятся центры афферентной иннервации различных мимических мышц лица, слизистой оболочки ротовой полости, глаза, лабиринта внутреннего уха и многих внутренних органов — органов дыхания, пищеварения, кровообращения. Спинномозговые афферентные нервы связывают продолговатый мозг с рецепторами кожи и скелетной мускулатуры. Сфера рецепции, на которую опирается в своей рефлекторной деятельности продолговатый мозг, значительно расширена по сравнению со спинномозговой рецепцией.
Эфферентные волокна черепно-мозговых нервов, равно как связанные с продолговатым мозгом эфферентные волокна спинномозговых путей, в том числе и вегетативной нервной системы, формируют рефлекторные дуги, при помощи которых продолговатый мозг осуществляет рефлекторную иннервацию жизненно важных органов.
Основная биологическая роль рефлекторной деятельности продолговатого мозга заключается в обеспечении через регуляцию функций иннервируемых им органов постоянства внутренней среды организма (гомеостаза). На уровне продолговатого мозга осуществляется рефлекторная регуляция таких жизненно важных функций, как акт дыхания, защитные рефлексы, связанные с деятельностью дыхательной системы (чихание, кашель), регуляция сердечно сосудистой деятельности, деятельности пищеварительного аппарата (как секреторной, так и моторной), рефлексы сосания, жевания, глотания, рвоты, моргания, слезотечения, потоотделения и т.п.).
Наряду с регуляцией вегетативных функций продолговатый мозг за счет связи с проприорецепторами выполняет роль регулятора тонуса скелетной мускулатуры. Рефлекторная деятельность продолговатого мозга обеспечивает тоническое напряжение прежде всего разгибателей, предназначенных для преодоления силы тяжести (позотонические рефлексы, осуществляющиеся при участии лабиринтных рецепторов и проприорецепторов скелетных мышц).
Наряду с рефлекторной и тонической функциями, продолговатый мозг выполняет и проводниковую функцию. Достаточно указать, что через него осуществляется эфферентная нисходящая связь между двигательными зонами коры больших полушарий и двигательными центрами спинного мозга (пирамидный тракт). Через продолговатый мозг осуществляется и афферентная связь между спинным мозгом и выше лежащими отделами.
К указанному можно прибавить, что продолговатый мозг как относительно высший отдел ЦНС регулирует работу более примитивного спинного мозга. Эта координационная функция нацелена на объединение всех сегментов спинного мозга в единое целое, на обеспечение условий для целостной деятельности спинного мозга.
Таблица 1. Центры и рефлексы продолговатого мозга.
| Центры продолговатого мозга | Рефлексы продолговатого мозга |
| 1) Дыхательный | Жизненно-важные рефлексы |
| 2) Сердечно-сосудистый | Защитные рефлексы |
| 3) Слюноотделения | Рефлексы пищевого поведения |
| 4) Слезоотделения | Рефлексы поддержания позы |
| 5) Кашля | Вегетативные рефлексы |
| 6) Чихания | Вестибуловегетативные |
| 7) Мигания | |
| 8) Рвоты | |
| 9) Сосания | |
| 10) Жевания | |
| 11) Глотания | |
| 12) Поддержания позы |
Повреждение продолговатого мозга связано с нарушением или полным выпадением функций, регулируемых этим отделом ЦНС. Очевидно, что полное разрушение продолговатого мозга ведет к гибели животных, для которых относительное постоянство внутренней среды является обязательным условием существования. Более примитивные животные переносят в течение известного времени разрушение продолговатого мозга, но и у них рефлекторная регуляция функций претерпевает известные нарушения. У человека заболевания продолговатого мозга чрезвычайно тяжело сказывается на всех видах деятельности организма. Непосредственной причиной неизбежной смерти является нарушение деятельности дыхательной системы.
Понравилась статья? Добавь ее в закладку (CTRL+D) и не забудь поделиться с друзьями:
Источник