Конечный мозг
Конечный мозг—telencephalon (рис. 159 и 160—1, 9)—часть головного мозга, которая чрезвычайно сильно прогрессирует в ряду животных и наивысшего своего развития достигает в классе млекопитающих, в особенности у приматов. У человека конечный мозг настолько перерастает все остальные отделы мозга, что почти все их прикрывает. Конечный мозг разделяется на обонятельный мозг—rhinencephalon—и плащ—pallium.
Обонятельный мозг
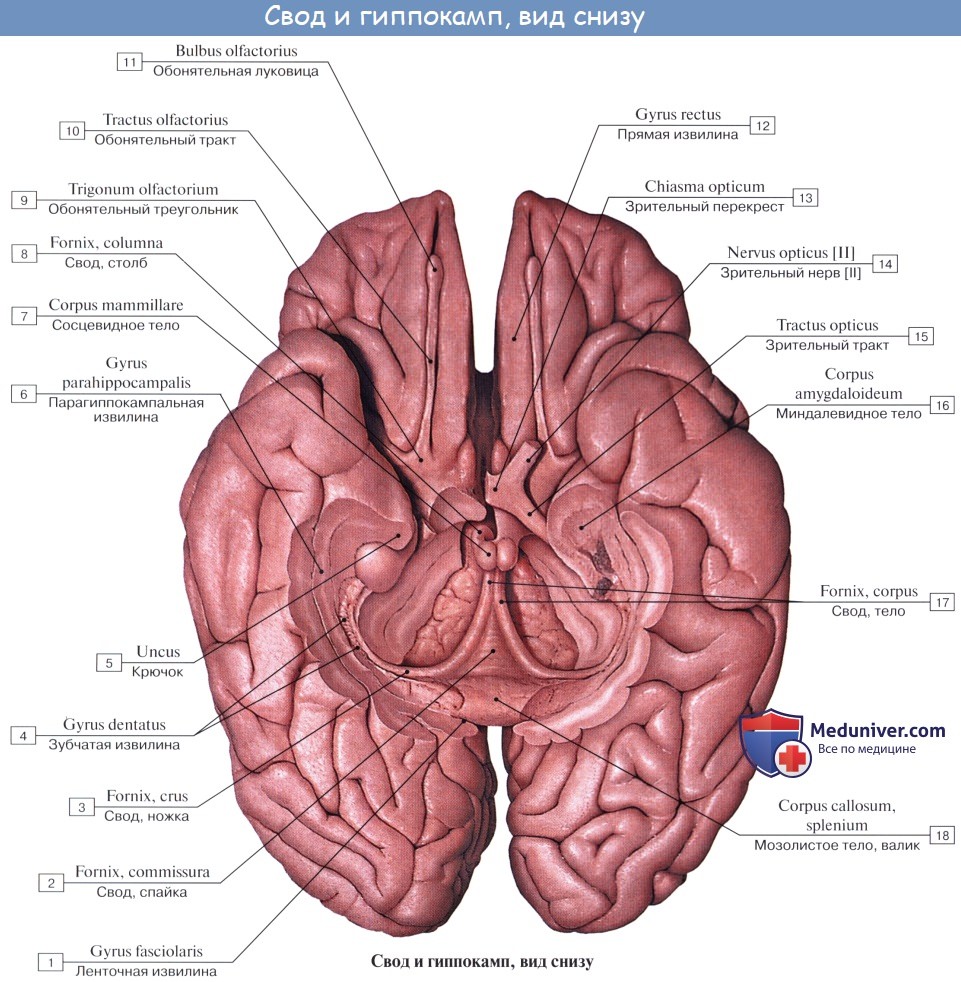
Из передней стенки конечного мозга разрастается парное выпячивание—обонятельные луковицы—bulbi olfactorii (рис. 159—10),—соединяющиеся с мозгом широким основанием или более или менее длинными ножками—обонятельными трактами—tractus olfactorius (у костистых рыб и у рептилий). Основание, или базальная часть, мозга принимает значительные размеры уже в примитивном мозге. Оно служит вторичным обонятельным центром и образовано значительным утолщением— базальным ганглием—nucleus basalis (1),—из которого у млекопитающих происходит полосатое тело—corpus striatum,—вдающееся в полость боковых желудочков мозга. Серое мозговое вещество полосатого тела разделяется в процессе развития белым мозговым веществом, формирующим внутреннюю капсулу—capsula interna (рис. 184—9),—на дорзальную часть, или хвостатое тело—nucleus caudatus,—и вентральную часть, или чечевицеобразное ядро (2)—nucleus lentiformis,—а наружная капсула—capsula externa (4)— отделяет латеральную часть,или ограду (3),—claustrum. Вследствие сильного разрастания полосатые тела, вдавливаясь в полость боковых желудочков, превращают их в щелевидные пространства (5). В высокодиференцированном мозге млекопитающих обонятельная часть его развита в ряду животных далеко не одинаково. У макросматических животных (к ним относятся все домашние животные и вообще большинство животных, обладающих хорошо развитым чувством обоняния) обонятельная часть мозга довольно значительная. У микросматических животных (обезьяны, зубастые киты—дельфины, кашалоты) соответственно функционированию органа обоняния и обонятельная часть мозга невелика. Из отдельных частей обонятельного мозга особенно выделяются обонятельные луковицы, которые у макросматических животных являются придатками мозга, выступающими спереди (рис. 177—I), а у микросматических животных они очень малы. Наружная (вентральная) часть базального ганглия, или обонятельная кора, формирует впереди перекрёста зрительных нервов, между латеральным и медиальным обонятельными трактами, обонятельный бугор—tuberculum olfactorium,—который среди домашних животных наиболее резко выражен только у свиней; у остальных он плоский, вследствие чего и называется просто обонятельным треугольником. У микросматических животных соответствующий участок едвa выступает.
Строение коры большого мозга
Вторичный плащ построен из серого и белого мозгового вещества. Первое располагается поверхностно и образует кору большого мозга, второе находится в глубине. Кора головного мозга в различных её участках гистологически устроена не одинаково, что впервые было установлено В. А. Бецом (1874) и им же объяснено: «Эта структурная диференциация мозговой коры есть выражение локализированности функций в коре» (1881). Различие касается как расположения клеточных элементов—цитоархитектоники, так и хода волокон в сером мозговом веществе—миэлоархитектоники. Цито- и миэлоархитектоника коры большого мозга более или менее подробно изучены лишь у человека и довольно слабо у животных. Клеточные элементы коры располагаются параллельно поверхности мозга в шесть слоёв (пластов). Пласты эти, считая снаружи внутрь, следующие: I—молекулярный, II—наружный зернистый, III—слой малых пирамидных клеток, IV—внутренний зернистый, V—слой больших пирамидных клеток и VI—слой полиморфных клеток (рис. 166). В филогенетическом ряду животных не все шесть слоёв бывают резко разграничены, но наличие их можно доказать у всех млекопитающих.

Клетки, образующие кору, делятся на три типа: Рамон-Кахалевские, пирамидные и полиморфные. Из них особого внимания заслуживают пирамидные клетки—клетки Беца: чем выше по своей организации или старше животное, тем больше у них отростков, следовательно, и связи этих клеток с другими обширнее (рис. 167). Различным слоям клеток приписывают различные функции: внутреннему зернистому слою (IV), являющемуся первичным, принадлежит рецепторная функция, клеткам V и VI слоёв, большим пирамидным и веретенообразным клеткам—эффекторная. Позже появляющиеся клетки II и III слоёв выполняют ассоциационные функции высшего порядка, т. е. психические. На основе местных различий в цитоархитектонике кору млекопитающих животных разделяют на главные области—поля—area,—причём каждое поле, по крайней мере у высших млекопитающих и особенно у человека, на основе различной миэлоархитектоники может быть подразделено ещё на ряд миэлоархитектонических участков. Филогенетически все поля во вторичном плаще диференцируются из примитивных четырёх областей низших млекопитающих (сумчатых и насекомоядных); эти области различны по своей цитоархитектонике, и, судя по наличию в них определённых проекционных волокон (рис. 165), им также соответствуют специальные функциональные центры. Так, в лобной области лежит примитивный моторный корковый центр, в затылочной—зрительный пентр, между ними—кожно-чувствующий, в височной области—слуховой пентр. Физиологические и архитектонические центры не всегда соответствуют расположению извилин. Соотношение мякотных волокон в коре мозга до известной степени пределяется принятым делением их на группы соответственно шести слоям клеток (рис. 168, справа).

По развитию миэлиновой оболочки (миэлогенетически) все нервные волокна относят или к проекционной системе, или ассоциационной. Первые развиваются уже до рождения, вторые—только после рождения. Миэлогенетически кора также может быть разделена по двум признакам—процентному соотношению проекционных и ассоциационных волокон и их онтогенезу—на области: первичную, конечную и промежуточную, т. е. волокна, одевающиеся миэлиновой оболочкой до рождения, после рождения b в последний период эмбрионального состояния. В настоящее время, на основании изучения цито- и миэлоархитектоники коры мозга, у человека выделяют более 250 полей. Ассоциационные волокна соединяют друг с другом многочисленные участки коры внутри одного полушария. Они делятся на короткие и длинные; первые связывают соседние извилины, вторые—более удалённые друг от друга участки и доли. Длинные волокна особенно сильно развиты у человека, в то время как у животных они или незначительны, или совсем отсутствуют. Коммиссуральные волокна соединяют участки, лежащие в разных полушариях. Они образуют переднюю и заднюю коммиссуры (спайки)—спайку аммонова рога и мозолистое тело. Проекционные волокна анатомически делятся на короткие и длинные, а функционально—на кортико-фугальные и кортико-петальные. Короткие проекционные волокна соединяют кору с ядрами зрительных бугров и четверохолмия. Длинные пути связывают кору с ядрами моста или со спинным мозгом. Все эти пути, как короткие, так и длинные, проходят между хвостатым телом и ядрами зрительных бугров, с одной стороны, и чечевичным ядром, с другой, образуя внутреннюю капсулу—capsula interna (рис. 184—9). На среднем мозге эти волокна формируют ножки большого мозга—pedunculi cerebri. Проекционные пути, так же как и ассоциационные, наиболее сильноразвиты у человека; у животных некоторые из них даже совсем отсутствуют. Таким образом, полушария большого мозга в целом и кора полушарий в частности устроены наиболее сложно и притом не одинаково в ряду млекопитающих, достигая максимума своего развития у высших приматов и особенно у человека. По И. П. Павлову, на большие полушария беспрерывно падают бесчисленные раздражения как из внешнего мира, так и из внутренней среды самого организма. Так как кора мозга высших животных является носительницей замыкательной функции (т. е. функции приобретения, образования новых связей между организмом и средой, развития нового жизненного опыта, функции онтогенетической адаптации, приспособляющей организм к условиям среды), высший отдел центральной нервной системы держит в своём ведении все явления, происходящие в теле.
Источник
Конечный мозг животного анатомия
Конечный мозг, telencephalon. Полушария, hemispheria cerebri
Как уже отмечалось, конечный мозг, telencephalon, представлен двумя полушариями, hemispheria cerebri. В состав каждого полушария входят: плащ, или мантия, pallium, обонятельный мозг, rhinencephalon, и базальные ядра.
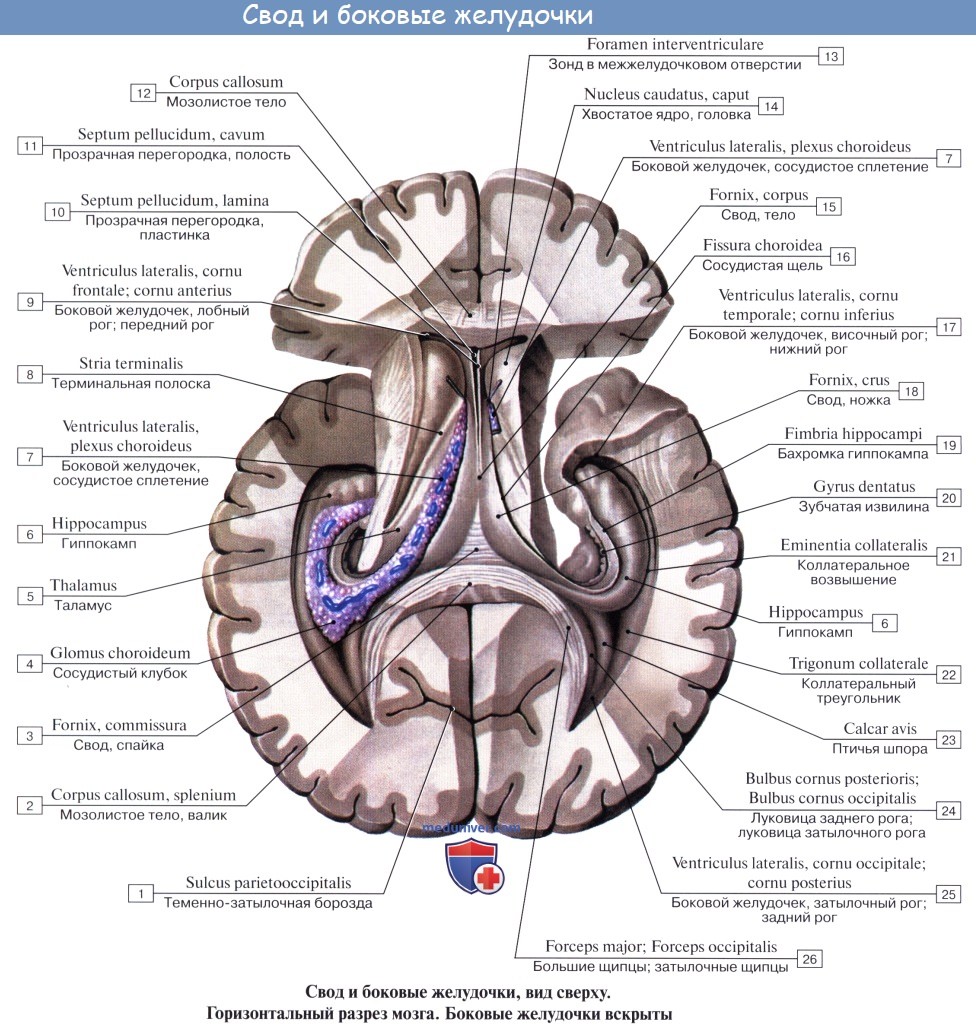
Остатком первоначальных полостей обоих пузырей конечного мозга являются боковые желудочки, ventriculi laterales. Передний мозг, из которого выделяется конечный, вначале возникает в связи с обонятельным рецептором (обонятельный мозг), а затем он становится органом управления поведением животного, причем в нем возникают центры инстинктивного поведения, основанного на видовых реакциях (безусловные рефлексы), — подкорковые ядра и центры индивидуального поведения, основанного на индивидуальном опыте (условные рефлексы), — кора большого мозга.
Соответственно этому в конечном мозге различают в порядке исторического развития следующие группы центров:
1. Обонятельный мозг, rhinencephalon, — самая древняя и вместе с тем самая меньшая часть, расположенная вентрально.
2. Базальные, или центральные, ядра полушарий, «подкорка», — старая часть конечного мозга, paleencephalon, скрытая в глубине.
3. Серое вещество коры, cortex, — самая молодая часть, neencephalon, и вместе с тем самая большая часть, покрывающая остальные как бы плащом, откуда и ее название «плащ», или мантия, pallium.
Кроме отмеченных для животных двух форм поведения, у человека возникает третья форма — коллективное поведение, основанное на опыте человеческого коллектива, создающегося в процессе трудовой деятельности человека и общения людей с помощью речи. Эта форма поведения связана с развитием самых молодых поверхностных слоев мозговой коры, составляющих материальный субстрат так называемой второй сигнальной (словесной) системы действительности (И. П. Павлов).
Так как в процессе эволюции из всех отделов центральной нервной системы быстрее и сильнее всего растет конечный мозг, то он у человека становится самой большой частью головного мозга и приобретает вид двух объемистых полушарий — правого и левого, hemispheria dextrum et sinistrum.
Учебное видео анатомии конечного мозга — обонятельного, базальных ядер, коры
Редактор: Искандер Милевски. Дата последнего обновления публикации: 22.7.2021
Источник