Обзор строения почек у позвоночных
Среди круглоротых у некоторых миксин вытянутая вдоль всего тела узкая почка содержит всего по одному нефрону на сегмент, очень короткие канальцы и потому считается пронефрической. Нужно иметь в виду, что функции выделительной системы у миксин упрощены – это единственные представители позвоночных, не поддерживающие активно осмотического равновесия с внешней средой – осмоконформеры, в отличие от осморегуляторов. У миноги имеется по нескольку нефронов на сегмент, что дает основания относить ее почку к мезонефрической. Но у нее сохраняется яркий примитивный признак: ее клубок артериол не подразделен, он остается общим для всех нефронов почки и тянется тонким жгутом на протяжении всей почки. Примыкающие к нему боуменовы капсулы разделены неполностью, будучи тесно упакованы бок обок одна с другой. Мочеточник тянется вдоль всей почки по ее вентральному краю.
Почку хрящевых рыб, как и у всех остальных анамний, считают мезонефрической. Соль выводится в обход почки – через ректальную железу, которая открывается в задний отдел кишечника. Длинный передний отрезок почки теряет выделительную функцию: у самца его небольшой фрагмент участвует в выведении мужских половых продуктов в качестве посредника между семенником и семяпроводом (вольфовым каналом), а у самки тот и другой просто редуцированы. Задняя часть почки усиленно развита в качестве органа выделения, но обслуживается не вольфовым каналом, а самостоятельным «вторичным» мочеточником, который развивается как отросток конечного участка вольфова канала. У самок некоторых видов редукция передней части почки выражена заметно слабее, чем у самцов.
У пресноводных костных рыб, ткани которых осмотически насасывают воду из внешней среды, главная задача почки — выведение этой избыточной воды путем фильтрации и последующей реабсорбции ценных веществ из первичной мочи. Главные компоненты нефрона — соответственно почечное тельце и проксимальный извитой каналец. Удаление продуктов азотистого обмена происходит отчасти в обход почек – аммиак просто вымывается через жабры. У относительно примитивных костных рыб (двоякодышащих, осетровых, костных ганоидов) часть почки и вольфов канал принимают участие в выведении мужских половых продуктов, но архинефрический проток совмещает функции мочеточника и семяпровода, не обнаруживая у самки тенденции к редукции (как и передняя часть почки); добавочных мочеточников не возникает.
У костистых рыб почка не участвует в выведении половых продуктов и не испытывает сколько-нибудь заметного влияния со стороны репродуктивной системы.
У обитателей соленых вод нефроны построены иначе. У морских костистых рыб, осмотически теряющих воду из-за отсоса ее гипертонической внешней средой и пьющих морскую воду для компенсации этих потерь, ультрафильтрация, а вместе с ней и реабсорбция не так важны, как секреция гипертонического раствора, которая имеет место в дистальном извитом канальце; у некоторых рыб (например, у морского конька и других пучкожаберных) эта секреция исчерпывает функцию выделения (нет ни почечных телец, ни реабсорбционных отделов канальцев – это так называемая агломерулярная почка). Особый случай среди морских костных рыб составляет единственная современная кистеперая рыба, целакант латимерия. У нее, как и у акул, достигнуто осмотическое равновесие жидкостей тела с морской водой за счет накопления мочевины.
У самцов бесхвостых амфибий вольфов канал и мезонефрическая почка совмещают функцию выделения с выведением половых продуктов. У самцов хвостатых амфибий обслуживание семенников передними частями почек вытесняет функцию выделения. Передняя часть почки перестраивается точно так же, как у пластиножаберных рыб, вольфов канал специализируется в качестве семяпровода; при задней, функционирующей части почки развивается значительное число дополнительных мочеточников. У самок эти изменения выражены, как правило, слабее, часть почки, сохраняющая выделительную функцию, простирается вперед немного дальше.
У рептилий, почка которых традиционно считается метанефрической, как и у других амниот, имеется вторичный мочеточник, а вольфов канал у самцов всегда функционирует в роли семяпровода, получая мужские половые продукты из семенника через посредство придатка семенника (epididymis; производного лежащей на этом уровне эмбриональной почки — мезоненфроса).
Хотя для рептилий (прежде всего ящериц), как и для других амниот, известна способность очень сильно концентрировать мочу, абсорбируя из нее воду, петля Генле в нефронах у них отсутствует. Благодаря дополнительному коллоидному осмотическому давлению крови в нее диффундирует вода из первичной мочи; в результате ее концентрация поднимается, и малорастворимая мочевая кислота кристаллизуется, а значит — удаляется из раствора и не может повышать его осмотическое давление. По мочеточнику выводится жидкая белая кашица, которая дополнительно высушивается почти до твердого состояния в клоаке. Таким образом, способность ящериц сильно концентрировать мочу определяется низкой растворимостью главного азотсодержащего вещества — мочевой кислоты. Хорошо растворимые электролиты, которые могли бы помешать этому процессу, у рептилий активно реабсорбируются из первичной мочи в проксимальном извитом канальце, а из организма выводятся другим путем – через носовые железы, секретирующие гипертонический раствор.
У птиц имеет место близкая картина, хотя часть нефронов снабжена довольно короткой петлей Генле, позволяющей создавать повышенную концентрацию соли в интерстициальной ткани, окружающей канальцы, и тем самым высокий локальный градиент осмотического давления, отсасывающий воду из мочи.
У млекопитающих это свойство выражено еще сильнее: благодаря существованию в нефронах длинных петель Генле они концентрируют хорошо растворимую мочевину, но далеко не в той степени, в какой поддается концентрированию мочевая кислота. В связи с этим млекопитающие не нуждаются в использовании отдельного пути для удаления соли и выводят ее с мочой. Наиболее типична для млекопитающих бобовидная форма почки, на вогнутой стороне которой расположена расширенная начальная часть мочеточника – воронкообразная камера с перепончатыми стенками, так называемая лоханка. Если бобовидное тело почки рассечь на две «семядоли», на разрезе видны две зоны ее мякоти – корковое вещество, содержащее почечные тельца, проксимальные и дистальные извитые канальцы, и мозговое вещество, которое образует обращенные тупыми вершинами в лоханку пирамиды. Пирамиды сложены ориентированными вдоль радиусов кривизны тела почки петлями Генле и собирательными трубочками, открывающимися в лоханку.
А. Ромер (1992) предложил новый подход к классификации почек позвоночных.
Теоретически мыслимая идеальная примитивная почка, заслуживающая имени голонефрос, должна была содержать по одному нефрону на сегмент на всем протяжении тела и мочеточник, в который они все открывались. К состоянию голонефроса близки почки личинок миксин и гимнофион. Но в них специализированная передняя группа канальцев развивается первой и образует пронефрический проток. У анамний (кроме хрящевых рыб) она даже функционирует на ранних стадиях развития, а затем рано редуцируется. Это пронефрос. Всю остальную часть голонефроса А.Ромер рассматривает в качестве опистонефроса (т. е. задней почки); в этом понятии он объединяет все категории дефинитивной почки позвоночных, сохраняя термины мезонефрос и метанефрос лишь для обозначения различных эмбриональных почек амниот. У всех позвоночных, кроме миксин, строение голонефроса усложнено под действием двух факторов. Во-первых, в нем нарушена простая сегментальная структура (в каждом сегменте образуется множество нефронов, а в задней части туловища исходная сегментация нефротома вообще стерта), а во-вторых, имела место конкуренция почки с семенником за вольфов канал, которая протекала с переменным успехом в разных линиях эволюции и во многих случаях привела к формированию вторичных мочеточников.
Эмбриональная дифференциация гонад
Зачаток гонад лежит в нефрогонотоме медиальнее зачатков почки в виде половой складки. Она образована зачатковым (герминативным) эпителием, под которым лежит мезенхима.
У самца развитие получает мезенхимная часть, которая потом превращается в семенные трубочки семенника, в стенках которых идет сперматогенез; зачатковый эпителий у самца дегенерирует.
У самки первичные половые тяжи дегенерируют, а зачатковый эпителий гипертрофируется, в нем появляются так называемые вторичные половые тяжи, позже дающие начало фолликулам.
У круглоротых и костистых рыб гонады не имеют связи с почками. У костистых рыб они свернуты в мешковидные образования.
Источник
Петля генле во всех нефронах птиц
Выделение продуктов распада и регуляция водного обмена осуществляются преимущественно почками. Крупные (до 1-2% от массы тела) метанефрические почки лежат в углублениях тазового пояса. От каждой почки отходит мочеточник, открывающийся в клоаку. Мочевого пузыря у птиц нет.
У птиц, как и у большинства пресмыкающихся, в качестве выделяемого из организма конечного продукта белкового обмена образуется не мочевина, а мочевая кислота, легко выпадающая из раствора кристалликами, образующими белую кашицеобразную массу. Такая консистенция выделяемой мочи, видимо, обусловила редукцию мочевого пузыря у птиц. Размеры почечных клубочков в почках птиц относительно невелики, что снижает интенсивность фильтрации и уменьшает потери воды. Основная масса мочевой кислоты попадает в просвет почечных канальцев не столько путем фильтрации из клубочков, сколько путем секреции специальными железистыми клетками стенок почечных канальцев, оплетенных капиллярами воротной системы почек. Одновременно почечные канальцы (нефроны) выполняют и осморегуляторную функцию.
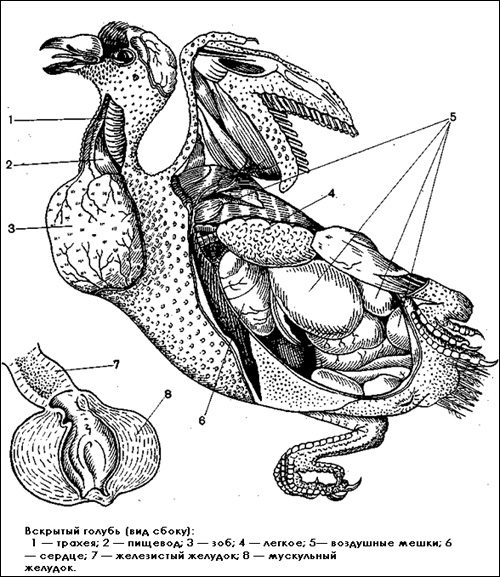
Вскрытый голубь — вид сбоку
В нефроне птиц появляется отсутствующий у анамний и у пресмыкающихся 11-образноизогнутый средний отдел — петля Генле, густо оплетенная капиллярами и функционирующая как своеобразная «умножительная противоточная система», использующая осмотический градиент — разное содержание солей в тканях почки и в просвете почечных канальцев. Проницаемость стенок петли Генле создает возможность пассивной реабсорбции воды из первичной мочи в зонад повышенной солености, а ионы натрия активно реабсорбируются в восходящей части петли и в концевой части канальцев. Этот механизм обеспечивает образование концентрированной мочи. Дополнительно всасывание воды проходит в клоаке. Все это позволяет удалять из организма продукты распада при минимальной потере воды.
У большинства птиц есть носовые железы, расположенные на лобных костях над орбитой. Особенно сильно они развиты у морских птиц (трубконосые, веслоногие и др.) и у некоторых пустынных птиц, которые вынуждены пить соленую воду. В секрете носовых желез, капельками стекающем через ноздри наружу, концентрация поваренной соли в 4-5 раз выше, чем в крови, и примерно вдвое выше, чем в морской воде. Выделение столь концентрированного раствора позволяет птицам пить воду соленых водоемов. Избыток других солей выделяется через почки с мочой.
Литература: Зоология позвоночных. Ч. 2. Пресмыкающиеся, птицы, млекопитающие. Наумов Н. П., Карташев Н. Н., Москва, 1979
Источник