Природа автоматии сердечной мышцы
Мембранная природа автоматии сердца. Водитель ритма. Пейсмекер. Проводимость миокарда. Истинный водитель ритма. Латентный водитель ритма.
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту же биоэлектрическую природу, что и в поперечно-полосатых мышцах. Наличие заряда на мембране здесь также обеспечивается разностью концентраций ионов калия и натрия возле ее внешней и внутренней поверхности и избирательной проницаемостью мембраны для этих ионов. В покое мембрана кардиомиоцитов проницаема для ионов калия и почти непроницаема для ионов натрия. В результате диффузии ионы калия выходят из клетки и создают положительный заряд на ее поверхности. Внутренняя сторона мембраны становится электроотрицательной по отношению к наружной.
В клетках атипического миокарда, обладающих автоматией, мембранный потенциал способен спонтанно уменьшаться до критического уровня, что приводит к генерации потенциала действия. В норме ритм сердечных сокращений задается всего несколькими наиболее возбудимыми клетками синоатриального узла, которые называются истинными водителями ритма, или пейсмекерными клетками. В этих клетках во время диастолы мембранный потенциал, достигнув максимального значения, соответствующего величине потенциала покоя (60—70 мВ), начинает постепенно снижаться. Этот процесс называют медленной спонтанной диастолической деполяризацией. Она продолжается до того момента, когда мембранный потенциал достигает критического уровня (40—50 мВ), после чего возникает потенциал действия.
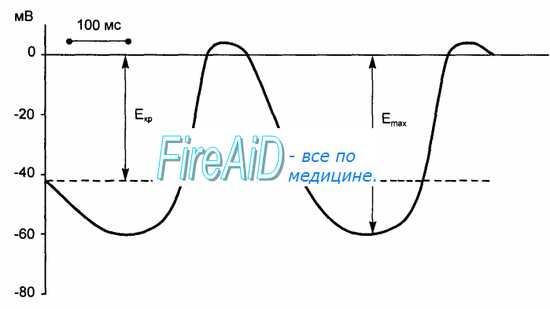
Рис. 9.6. Развитие потенциала действия истинного водителя ритма автоматии сердца. Во время диастолы спонтанная деполяризация уменьшает мембранный потенциал (Еmах) до критического уровня (Еkp) и вызывает потенциал действия.
Для потенциала действия пейсмекерных клеток синоатриального узла характерны малая крутизна подъема, отсутствие фазы ранней быстрой реполяризации, а также слабая выраженность «овершута» и фазы «плато». Медленная реполяризация плавно сменяется быстрой. Во время этой фазы мембранный потенциал достигает максимальной величины, после чего вновь возникает фаза медленной спонтанной деполяризации (рис. 9.6).
Частота возбуждения пейсмекерных клеток у человека составляет в покое 70—80 в минуту при амплитуде потенциала действия 70—80 мВ. Во всех остальных клетках проводящей системы потенциал действия в норме возникает под влиянием возбуждения, приходящего из синоатриального узла. Такие клетки называют латентными водителями ритма. Потенциал действия в них возникает раньше, чем их собственная медленная спонтанная диастолическая деполяризация достигает критического уровня. Латентные водители ритма принимают на себя ведущую функцию только при условии разобщения с синоатриальным узлом. Частота спонтанной деполяризации таких клеток у человека составляет 30—40 в минуту (рис. 9.7).
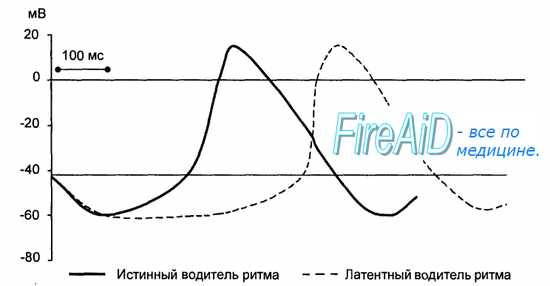
Рис. 9.7. Развитие потенциала действия истинного и латентного водителей ритма автоматии сердца. Скорость медленной диастолической деполяризации истинного водителя ритма больше, чем у латентного.
Спонтанная медленная диастолическая деполяризация обусловлена совокупностью ионных процессов, связанных с функциями плазматических мембран. Среди них ведущую роль играют медленное уменьшение калиевой и повышение натриевой и кальциевой проводимости мембраны во время диастолы, параллельно чему происходит падение активности электрогенного натриевого насоса. К началу диастолы проницаемость мембраны для калия на короткое время повышается, и мембранный потенциал покоя приближается к равновесному калиевому потенциалу, достигая максимального диастолического значения. Затем проницаемость мембраны для калия уменьшается, что и приводит к медленному снижению мембранного потенциала до критического уровня. Одновременное увеличение проницаемости мембраны для натрия и кальция приводит к поступлению этих ионов в клетку, что также способствует возникновению потенциала действия. Снижение активности электрогенного насоса дополнительно уменьшает выход натрия из клетки и, тем самым, облегчает деполяризацию мембраны и возникновение возбуждения.
Видео физиология фазы потенциала действия пейсмекерных клеток сердца
— Вернуться в оглавление раздела «Физиология человека.»
Источник
1. Автоматия сердца.
Автоматия сердца – это его способность к ритмическому сокращению без всяких видимых раздражений под влиянием импульсов, возникающих в самом органе. Автоматия сердечной мышцы бывает миогенной – когда импульсы появляются в самих мышечных волокнах, и нейрогенной – когда импульсы возникают в клетках нервных ганглиев. Миогенная автоматия обеспечивает сокращения сердца на ранних стадиях эмбрионального развития, а также некоторое время (несколько часов и даже суток) после перерезки всех идущих к сердцу нервов. В постэмбриональный период ритмическая деятельность сердца происходит благодаря наличию проводящей системы сердца. Так, в области ушка правого предсердия находится ведущий центр автоматизма — синусно-предсердного (синатриального) узла. Он является главным центром автоматии сердца – пейсмекером первого порядка. От него по рабочим клеткам миокарда и проводящим волокнам предсердий возбуждение достигает предсердно-желудочкового (атриовентрикулярного) узла, расположенного в стенке правого предсердия вблизи перегородки между предсердиями и желудочками. Этот узел является пейсмекером второго порядка. Далее возбуждение переходит на миокард желудочков по волокнам пучка Гиса (предсердно-желудочкового пучка) и достигает волокон Пуркинье (сердечных проводящих миоцитов). В обычных условиях частоту активности миокарда всего сердца в целом определяет синусно-предсердный узел. При нарушении автоматизма синусно-предсердного узла ритмические сокращения сердца могут продолжаться благодаря импульсам, возникающим в атриовентрикулярном узле. Однако частота и сила сокращений при этом вдвое меньше, чем до нарушений в области синусно-предсердного узла. В случае невозможности передачи возбуждения к желудочкам они начинают сокращаться в ритме пейсмекеров третьего порядка – клеток пучка Гиса и волокон Пуркинье. При повреждении всех водителей ритма сердце останавливается (искусственные кардиостимуляторы). Синусно-предсердный узел подчиняет себе все нижележащие образования проводящей системы, навязывая им свой ритм. Поэтому все отдельные части проводящей системы, хотя и имеют собственную активность, начинают работать в едином ритме. Явление, при котором структуры с замедленным ритмом генерации потенциалов усваивают более частый ритм других пейсмекерных участков называют усвоением ритма. Исходя из этого Гаскелл установил Закон градиента автоматизма сердца – у всех позвоночных степень автоматии тем выше, чем ближе расположен участок проводящей системы к синоатриальному узлу. Теории автоматизма. Существует несколько теорий, объясняющих происхождение автоматизма (нейрогенная, эндогенная и др.). Наиболее популярна теория диастолического поля, в соответствии с которой в начальную фазу диастолы в проводящих миоцитах регистрируется мембранный потенциал, равный -90 мВ. В диастолу метаболизм сердечной мышцы изменяется и МП постепенно уменьшается, постепенно достигая критического уровня деполяризации. Когда уровень потенциала покоя уменьшается по сравнению с исходным приблизительно на 2 мВ, наступает резкое увеличение проницаемости мембраны вначале для Na + , а позднее для Са 2+ . В результате этих процессов МП приближается к критическому уровню деполяризации, при достижении которого в клетках синусно-предсердного узла возникает ПД. Все остальные отделы сердца подчиняются возникшему ПД — возбуждению, генерируемому в водителе ритма.
Источник